



Genere Chrysellampus Semenov, 1932
Chrysellampus Semenov-Tian-Shanskij, 1932: 5. Type species: Ellampus heros Semenov, 1892, by original designation.
Parellampus Semenov-Tian-Shanskij, 1932: 7. Type species: Parellampus praeteritorum Semenov, 1932, by original designation. Syn. nov.
Chrysellampus Semenov: Semenov-Tian-Shanskij 1967: 119; Nikol'skaya 1978: 63 (part.).
Omalus (Chrysellampus) Panzer, 1801: Linsenmaier 1959: 22, 1997: 249 (part.), 1999: 22, 23 (part.); Martynova & Fateryga 2014: 14.
Omalus (Philoctetes) Panzer: Arens 2014: 570 (part.).
Systematics. The genus Chrysellampus Semenov, 1932 was traditionally considered as a valid genus (Tsuneki 1948, 1950, 1953a, 1953b; Semenov 1967; Nikol'skaya 1978) or valid subgenus (Linsenmaier 1959, 1997; Martynova & Fateryga 2014). Kimsey & Bohart (1991) proposed a new systematic classification of genera closely related to Omalus Panzer, 1801 and (1991: 251) synonymised Chrysellampus Semenov, 1932 with Philoctetes Abeille de Perrin, 1879. Later Linsenmaier (1997) revalidated Chrysellampus Semenov, 1932 as subgenus.
Description. Chrysellampus Semenov, 1932 was synonymised with Philoctetes Abeille de Perrin, 1879 by Kimsey & Bohart (1991), but it can be easily separated by the following characteristics: habitus subcylindrical and elongate (stocky and roundish in Philoctetes) (Martynova & Fateryga 2014: Fig. 14); F2–F11 flattened and dilated (Fig. 3) (cylindrical and not dilated in Philoctetes); MS bisected or nearly so by curved genal carina (Figs 2A, 3, 5, 8) (MS not bisected and genal carina not curved in Philoctetes); genal carina well developed and sharp from lower margin of temple to MS (Figs 3, 5, 8) (faint or not well developed in Philoctetes); pronotum with large and dense punctures (Figs 4A–4F, 6A–6F, 8) (smooth to sparsely punctate in Philoctetes); mesoscutum with large punctures clumped along notauli (C. sculpticollis and C. medanae) or covering entire surface (punctures only clumped along notauli or more evenly distributed, but anyway gathered together toward notauli in Philoctetes); if punctures on mesoscutum are clumped along notauli, then punctuation on pronotum is dense with large punctures (smooth to sparsely punctate in Philoctetes); metanotum round, with exception of C. praeteritorum (Semenov, 1932) (usually conical to mucronate in Philoctetes); medial vein weakly curved (strongly arched in Philoctetes); the distance between posterior margin of anterior declivity of T1 and posterior margin of T1 as long as or longer than mesoscutellum (Fig. 2C) (very short, 0.2–0.5 × as long as in Philoctetes (Fig. 2B)); apex of T3 with median notch, with tooth at each side (with or without median notch in Philoctetes); head and mesosoma characterized by colliculate sculpture (Harris 1979), a reticulate microsculpture continuously set with granulations on the intervals among punctures (Figs 1A, 1B) (polished in Philoctetes); tarsal claw with five teeth (two to four in Philoctetes).

FIGURE 1. Chrysellampus sculpticollis (Abeille de Perrin), ♀. A. Punctuation and colliculate sculpture of mesoscutum over tegula margin, 150x; B. Punctuation and colliculate sculpture of mesoscutum over tegula margin, close up, 300x; C. Left apical tooth with rim, 500x. (Agnoli & Rosa 2011).

FIGURE 2. A. Chrysellampus sculpticollis (Abeille de Perrin), ♂. Head, lateral view. Arrow pointing curved genal carina bisecting MS.
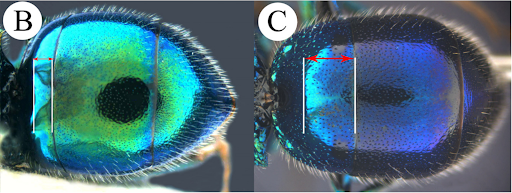
FIGURE 2. B. Philoctetes sp., metasoma, dorsal view. Arrow delimiting the distance between posterior margin of anterior declivity and posterior margin of T1; C. Chrysellampus sp., metasoma, dorsal view. Arrow delimiting the distance between posterior margin of anterior declivity and posterior margin of T1.
Species included. At present, the genus Chrysellampus Semenov includes ten rare and homogeneous species: C. sculpticollis (Abeille de Perrin, 1878), C. medanae (du Buysson in Magretti, 1890), C. heros (Semenov, 1892), C. pici (du Buysson, 1900) (=C. nigromaculatus Linsenmaier, 1997), C. harmandi (Buysson, 1903), C. praeteritorum (Semenov, 1932), comb. nov., C. duplipunctatus Tsuneki, 1948, C. tatianae Semenov, 1967, C. obtusidentibus Rosa, Wei & Xu, sp. nov., and C. proximocellis Rosa, Wei & Xu, sp. nov.
Biology. Chrysellampus are known as nest parasite of Crabronidae (Martynova & Fateryga 2014). Martynova & Fateryga (2014) gave description of larva and cocoon of C. sculpticollis, and its feeding habits and interactions with host, Psenulus fuscipennis (Dahlbom).
Distribution. Chrysellampus Semenov is known from Palaearctic and Oriental Regions (China: Yunnan).
From: Rosa P., Wei N.-S. & Xu Z.-F., 2015 - Revalidation of genus Chrysellampus Semenov, 1932, with description of two new species from China (Hymenoptera, Chrysididae). Zootaxa 4034 (1): 148–160.
European Species
Dichiarazione su copyright, autore e proprietà
Tutti i contenuti di questa pagina sono copyright ©️ Chrysis.net se non diversamente specificato - si prega di consultare i singoli casi per dettagli sulla paternità e sul copyright. Gli esemplari nelle foto provengono dalle collezioni personali degli autori o di altri collaboratori e dalle collezioni di vari musei. Salvo diversamente specificato, l'intero contenuto di questo sito Web è a scopo personale, non commerciale, scientifico ed educativo con giusto accredito alla pagina da cui è stato derivato, e conformemente ai Termini e condizioni di Chrysis.net.
Per citazioni
Agnoli G.L. & Rosa P. (2024) Risultati della ricerca , in: Chrysis.net website. Interim version 18 May 2024, URL: https://www.chrysis.net/it/?s=cash%2525252Baccounting%25252525E4%25252525BB%2525252580%25252525E9%25252525BA%25252525BC%25252525E6%2525252584%252525258F%25252525E6%2525252580%252525259D-%25252525E3%2525252580%2525252590%25252525E2%252525259C%2525252594%25252525EF%25252525B8%252525258F%25252525E6%252525258E%25252525A8%25252525E8%2525252596%25252525A6BB87%25252525C2%25252525B7CC%25252525E2%252525259C%2525252594%25252525EF%25252525B8%252525258F%25252525E3%2525252580%2525252591-%25252525E5%2525252582%25252525AC%25252525E7%252525259C%25252525A0%25252525E7%252525259C%25252525BC%25252525E9%252525258F%25252525A11%7E28%25252525E5%2525252585%25252525A8%25252525E7%25252525B3%25252525BB%25252525E5%2525252588%2525252597%25252525E4%25252525B8%252525258B%25252525E8%25252525BC%2525252589-cash%2525252Baccounting%25252525E4%25252525BB%2525252580%25252525E9%25252525BA%25252525BC%25252525E6%2525252584%252525258F%25252525E6%2525252580%252525259D547c2-%25252525E3%2525252580%2525252590%25252525E2%252525259C%2525252594%25252525EF%25252525B8%252525258F%25252525E6%252525258E%25252525A8%25252525E8%2525252596%25252525A6BB87%25252525C2%25252525B7CC%25252525E2%252525259C%2525252594%25252525EF%25252525B8%252525258F%25252525E3%2525252580%2525252591-%25252525E5%2525252582%25252525AC%25252525E7%252525259C%25252525A0%25252525E7%252525259C%25252525BC%25252525E9%252525258F%25252525A11%7E28%25252525E5%2525252585%25252525A8%25252525E7%25252525B3%25252525BB%25252525E5%2525252588%2525252597%25252525E4%25252525B8%252525258B%25252525E8%25252525BC%25252525893hnw-cash%2525252Baccounting%25252525E4%25252525BB%2525252580%25252525E9%25252525BA%25252525BC%25252525E6%2525252584%252525258F%25252525E6%2525252580%252525259Dta73d-%25252525E5%2525252582%25252525AC%25252525E7%252525259C%25252525A0%25252525E7%252525259C%25252525BC%25252525E9%252525258F%25252525A11%7E28%25252525E5%2525252585%25252525A8%25252525E7%25252525B3%25252525BB%25252525E5%2525252588%2525252597%25252525E4%25252525B8%252525258B%25252525E8%25252525BC%2525252589m10y%2Fpage%2F5%2Fin%2520internet%2Fpage%2F27%2Ftaxon.htm.